Productos finales de glicación avanzada (AGEs) y su importancia en enfermedades crónicas relacionadas con la nutrición
Marcela Fuentes1,a, Pablo Olmos1 y José Luis Santos1,b
Advanced glycation end products (AGEs) and its importance in chronic diseases related to nutrition
1Departamento de Nutrición, Diabetes y Metabolismo. Facultad de Medicina, Pontificia Universidad Católica de Chile, Santiago, Chile.
aDoctora en Nutrición.
bDoctor en Biología.
Correspondencia a:
Marcela Fuentes
E-mail: marcelafuentes@gmail.com
Recibido: 21-12-2014
Aceptado: 19-03-2015
Maillard reaction occurs when reducing sugars react in a non-enzymatic way with amino groups from proteins, lipids and nucleic acids. Products of this reaction are known as Advanced Glycation End Products (AGEs). These products are formed from endogenous sources (within the body) and exogenously (produced in food preparation, as well as those supported in their formation by tobacco smoke). In the food industry this reaction is known as “browning” and is directly related to cooking time of these, affecting its color and flavor. After food preparation and the formation of exogenous AGEs, these are absorbed in the digestive tract and are part of the pool of total body AGEs. AGEs alter structure and function of molecules and increase oxidative stress in biological systems. AGEs generally refers to non-reactive terminal products as CML (3,4-Ne-carboxymethyl-lysine), but also includes intermediate or precursor of AGEs as 3DG (3-deoxyglucosone), or MGO (methyl -glyoxal) and its derivatives. Glycation corresponds to a non-enzymatic glycosylation. This process contributes to protein post-translational modification. This process causes quantitative and qualitative changes in the extracellular matrix components which can affect cell adhesion, growth, and others. The process of protein glycation has been associated with development mechanisms of various diseases and complications such as retinopathy, nephropathy and neuropathy associated with diabetes, macrovascular disease, Alzheimer’s disease, cataracts, and aging.
Key words: AGEs, Chronic diseases, food nutrition.
En 1912 Louis Camille Maillard describió la reacción química que ocurre entre proteínas y azúcares reductores, y que forma un grupo heterogéneo de productos de color marrón y que se conoce como reacción de Maillard1. La reacción de Maillard ocurre cuando azúcares reductores reaccionan de una manera no-enzimática principalmente con grupos amino pertenecientes a proteínas, con lípidos y ácidos nucleicos. A los productos de esta reacción se les conoce como Productos Finales de Glicación Avanzada (Advanced glycation end products, AGEs). Ella ocurre tanto en el interior del organismo (es decir, de forma endógena) como también sucede en la preparación de alimentos que contienen azúcares, lípidos y proteínas y son procesados (cocción, fritura, deshidratación, etc.), además el humo del tabaco también da lugar a AGEs producidos de forma exógena2,3. La formación de AGEs in vivo e in vitro, es dependiente de la tasa de recambio de los blancos químicamente modificados, tiempo, y concentración de azúcar4. En la industria de los alimentos esta reacción es conocida como “pardeamiento” y está relacionada directamente al tiempo de cocción de estos, influyendo en su color y sabor5. Posterior a la preparación de los alimentos y la formación de AGEs exógenos, estos son absorbidos en el tubo digestivo y forman parte del pool de AGEs totales del organismo. Los AGEs alteran la estructura y función de moléculas e incrementan el estrés oxidativo en los sistemas biológicos2.
En general el término AGEs hace referencia a productos terminales no-reactivos tal como CML (3,4-Ne-carboximetil-lisina), aunque también incluye a intermediarios o precursores de AGEs como 3DG (3-deoxiglucosona), o MG (metil-glioxal) y sus derivados. Entre los azúcares presentes naturalmente, la glucosa presenta la tasa de glicación más lenta, mientras que azúcares intracelulares como fructosa, treosa, glucosa-6-fosfato y gliceraldehído- 3-fosfato, forman AGEs de forma mucho más rápida6-8.
Es necesario hacer notar, que la glicación corresponde a una glicosilación no-enzimática. Este proceso contribuye a la modificación post-traduccional de proteínas9. Intracelularmente el impacto de la glicación es contrarrestado por un alto recambio y corta vida-media de muchas proteínas celulares. Sin embargo, extracelularmente, proteínas de larga vida-media acumulan productos de glicación de forma significativa con el tiempo10,11. Este proceso de glicación no-enzimática de proteínas causa cambios cuantitativos y cualitativos en los componentes de matriz extracelular12-16 lo cual puede afectar la adhesión celular, crecimiento, y producción de ésta17-19.
Formación de AGEs
En la Figura 1 podemos ver un esquema general y resumido en el que se muestran los que llevan a la generación de AGEs. La formación de AGEs ocurre a través de una serie de reacciones químicas; en la primera de ellas, el grupo carbonilo de una cetona o aldehído de un azúcar reductor, se une a un aminoácido libre (principalmente lisina y arginina) de una proteína, lípido o DNA, de una manera no-enzimática para formar una base de Schiff. Los aductos tempranos de glicación (Early glycation adducts, EGAs) corresponden a estos productos iniciales (bases de Schiff y productos de Amadori, o fructosaminas). Los mecanismos de reacción que llevan desde EGAs hasta AGEs, no han sido completamente dilucidados, aunque han sido ampliamente estudiados10. El inicio de estas transformaciones depende de la concentración de glucosa y tiene lugar dentro de unas pocas horas. Si la concentración de glucosa disminuye, la reacción es aún reversible. Sin embargo, si la reacción sigue su curso, la base de Schiff sufre un reordenamiento químico y forma los llamados productos de Amadori o fructosamina, esto ocurre en un período de días5,20,21. Los productos de Amadori son más estables que las bases de Schiff, pero aún en este punto la reacción es parcialmente reversible. Un ejemplo de producto de Amadori es la hemoglobina glicosilada (HbA1c), la cual se forma por un residuo de valina N-terminal de una cadena de b hemoglobina que reacciona con glucosa y que es usualmente usada como un marcador del control glicémico. Si estas reacciones se continúan desarrollando y la hiperglicemia persiste, se llegará a una acumulación de productos de Amadori, los cuales sufrirán complejos reordenamientos químicos (oxidaciones, reducciones e hidrataciones) y formarán proteínas entrecruzadas20,22.
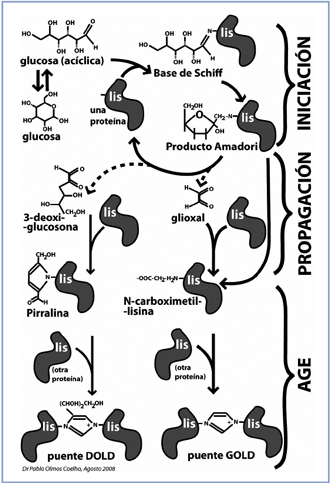 Figura 1. Formación de AGEs. (AGEs: Advanced glycation end products, Productos
finales de glicación avanzada, MG: metil-glioxal, 3DG: 3-deoxiglucosona,
CML: carboxi metil-lisina). Figura facilitada por Dr. Pablo Olmos36.
Figura 1. Formación de AGEs. (AGEs: Advanced glycation end products, Productos
finales de glicación avanzada, MG: metil-glioxal, 3DG: 3-deoxiglucosona,
CML: carboxi metil-lisina). Figura facilitada por Dr. Pablo Olmos36.
Posteriormente, se forman los dicarbonilos oxidantes glioxal y 3-deoxiglucosona, que son producto de la desglicosilación de parte del producto de Amadori, y que son potentes agentes alicantes y oxidantes, capaces de catalizar nuevas reacciones de unión de glucosa a proteínas. En esta fase ocurren varias reacciones de glico-oxidación proteica, todas ellas tendientes a formar productos de glicación que, como están unidos a una sola proteína, no forman puente entre dos de ellas (pirralina y N-carboxi metil-lisina)22,23. Este proceso se desarrolla a lo largo de semanas, e incluso meses, y es irreversible. Sin embargo, a pesar de la lentitud de la formación de estas estructuras, su formación puede ser catalizada por la presencia de estrés oxidativo, la auto-oxidación de la glucosa, la peroxidación lipídica, la presencia de iones metálicos y otros catalizadores, los cuales pueden aumentar sustancialmente la formación post-Amadori de AGEs10,24-27.
La fase final en la formación de AGEs comienza con la unión de la pirralina y de la N-carboximetil-lisina con una segunda proteína, formando estructuras conocidas como puentes DOLD y GOLD, los cuales alteran irreversiblemente las estructuras terciarias y cuaternarias de las proteínas22,23,28,29. Estas estructuras no tienen propiedades fluorescentes, pero hay otros AGEs que sí fluorescen, como las estructuras de “puente glucoespano” y “puente pentosidina”. Todos estos AGEs son muy estables a fuerzas mecánicas y degradación proteolítica, debido a las estructuras entrecruzadas que se forman durante la glicación30 y se acumulan dentro y fuera de las células e interfieren con la función de las proteínas27,28.
La glicación de proteínas se acompaña de un incremento en la actividad de los radicales libres28,31-33. No sólo es relevante la aparición de estas moléculas, sino que su interacción con sus propios receptores, los cuales son conocidos como RAGE.
Los AGEs pueden ser medidos y detectados por el uso de anticuerpos a través de inmunoensayos comerciales, inmunohistoquímica o medición de fluorescencia. Ya mencionamos que la HbA1c es un ejemplo de producto de Amadori que puede ser medido y servir también como indicador del control glicémico. Por su parte, la medición del contenido de AGEs por fluorescencia en biopsias de piel es un indicador más preciso que HbA1c para predecir la incidencia y progresión de complicaciones diabéticas a 10 años34. La American Diabetes Association describe la determinación de HbA1c como uno de los criterios diagnósticos para examinar la presencia de diabetes o pre-diabetes, y establece un valor normal menor a 5,7%35.
Receptores de AGEs
Los receptores específicos de AGEs son capaces de modular la captación y remoción de AGEs desde las células, a través de la endocitosis y degradación de moléculas modificadas de AGEs.
Los RAGE son miembros de la superfamilia de las moléculas de superficie celular de tipo inmunoglobulina, los cuales son capaces de reconocer un amplio rango de estructuras químicas y son expresados en una gran variedad de tipos celulares37,38. Se componen de una región extracelular que contiene un dominio inmunoglobulina tipo-V y dos dominios tipo-C38. Hay tres grandes grupos de RAGE, los cuales son variantes de splicing: RAGE de tamaño normal y completo, RAGE truncado en N-terminal y RAGE truncado en C-terminal, también conocido como es RAGE (endogenous secretory RAGE), la cual posee todos los dominios extracelulares, pero carece de los de transmembrana e intracitoplasmáticos39,4. Las formas solubles de RAGE finalmente neutralizan las acciones de AGEs sobre RAGE de superficie celular.
Los RAGE son inducidos por señales pro-inflamatorias, y su actividad biológica es dependiente de su unión a una variedad de ligandos41,42. Los RAGE actúan como un receptor de diversos ligandos liberados por células inflamadas, estresadas y dañadas. La expresión aumentada de RAGE de superficie celular y la acumulación de sus ligandos, ha sido observada en un amplio rango de desórdenes caracterizados por inflamación crónica, tales como la enfermedad intestinal crónica, artritis reumatoide, aterosclerosis, enfermedad de Alzheimer, y las complicaciones vasculares de la diabetes43. Las respuestas desencadenadas por la interacción ligando-receptor son muy amplias, e incluyen principalmente activación de las siguientes vías de señalización post-receptor de estas familias de factores de transcripción: NF-kB, “factor nuclear -kB”44, CREB, “cAMP response element-binding”45, EGR-1, “Early growth response protein 1”46 y AP-1, “Activator protein-1”47. Además, RAGEs activan un rango muy amplio de cascadas de transducción de señales: la familia de las proteínas quinasas activadas por mitógeno (MAP-quinasas), miembros de la familia de señalización JAK-STAT, CDC42, RAC1 y otros miembros de la familia Ras, SRC1, miembros de la familia de señalización SMAD, y fosfatidil-inositol3-quinasa47-52.
Otros receptores de AGE, son AGE-R1 (también conocida como p60), AGE-R2 (también llamada p90) y AGE-R3 (también conocida como Galectina-3, Mac-2 o Proteína de unión-35)53-55. Estos receptores pueden ser regulados por factores tales como la glucosa, AGEs, especies reactivas del oxígeno, entre otros56.
AGEs en los alimentos
Los AGEs están presentes en una amplia variedad de alimentos, preferentemente en aquellos procesados. Hay un gran número de estudios que muestran la asociación entre la dieta y AGEs, referidos principalmente al procesamiento de los nutrientes y el efecto que este tiene en la formación de AGEs. Se ha analizado y medido qué factores producen un aumento en la formación de AGEs57. El calor aplicado en la cocción está directamente asociado al aumento en AGEs y otros compuestos dañinos para la salud, como por ejemplo aminas heterocíclicas y acrilamida. Un aspecto interesante, es que en ausencia de proteínas o calor, los niveles de AGEs no correlacionan con contenidos altos de azúcar, ni tampoco que la ausencia de azúcar predice bajos niveles de AGEs (como en preparaciones que contienen aditivos de caramelo preformados similares a AGEs). En la Figura 2, podemos apreciar algunos alimentos de consumo habitual, y sus concentraciones de CML.
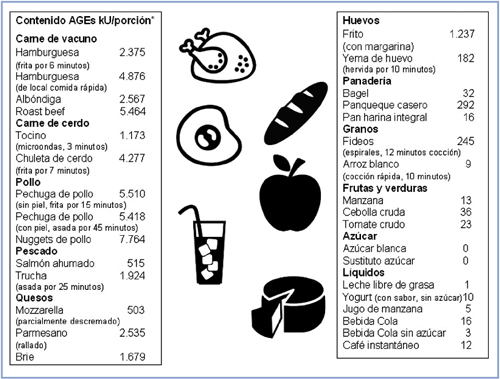 Figura 2. Concentración de AGEs (kU/porción) en función de mediciones de CML por la técnica de ELISA. Adaptado de Uribarri et al (2010)58.
Figura 2. Concentración de AGEs (kU/porción) en función de mediciones de CML por la técnica de ELISA. Adaptado de Uribarri et al (2010)58.
En general, y basándose en la ecuación:
Elevación en suero de AGEs (mg/dL) x volumen de plasma (dL) = Cantidad de AGEs en sangre (mg) que han sido absorbidos oralmente resultó ser aproximadamente el 10% de cantidad estimada a estar presente en el alimento ingerido. De los cuales, sólo un tercio fue excretado en la orina de personas con función renal normal57.
La evidencia de que la concentración de AGEs se ve incrementada en el suero y la orina de individuos normales, después de ingerir una dieta rica en AGEs, confirma que las estructuras conservadas en los AGEs sobreviven al proceso digestivo y que son transportados como moléculas de bajo peso molecular por el torrente sanguíneo, en conjunto con péptidos cortos y aminoácidos presentes en la digestión, en una manera directamente proporcional a la cantidad ingerida57. Estos productos son consumidos de forma sostenida y acumulativa en la dieta, por años. Cabe hacer notar que las dietas de los niños son procesadas de la misma forma. Por otra parte, hay que mencionar, que la calidad nutricional de los alimentos se ve afectada durante el procesamiento, principalmente porque el calor induce la pérdida de aminoácidos esenciales durante reacciones de entrecruzamiento entre ellos, o durante reacciones de tipo Maillard con carbohidratos reductores.
Excreción de AGEs
El principal mecanismo de degradación de tejidos y células modificados por AGE es a través de receptores específicos de AGE en macrófagos59. Después de la degradación, pequeños péptidos solubles de AGE son liberados y depurados por los riñones. Un deterioro en la función renal se traduce en una acumulación de AGE que puede llevar a una perturbación endotelial y, por lo tanto, a una enfermedad vascular4. En este contexto, el “clearance” urinario de AGEs correlaciona directamente con el “clearance” de creatinina (Ccr)59,60.
En individuos diabéticos, o con enfermedad renal, sin embargo, la excreción renal de AGEs se ve afectada, por lo que presentan niveles elevados de AGEs en suero y excreción urinaria de AGEs reducida59,60. Esta situación implica que los AGEs se acumulan plasmáticamente y por lo tanto, está el riesgo cierto de que reaccionen y se produzcan nuevos entrecruzamientos con proteínas plasmáticas60,61.
Sólo un tercio de los AGEs detectados en el suero son detectados en la orina después de 48 h62,63, el resto probablemente se une covalentemente en tejidos y células57.
Patologías asociadas a AGEs
El proceso de glicación de proteínas se ha asociado con mecanismos de desarrollo de diversas enfermedades y complicaciones, como retinopatía, neuropatía y nefropatía asociadas a diabetes mellitus64, enfermedad macrovascular65, enfermedad de Alzheimer66, cataratas11 y envejecimiento10.
Las complicaciones de la diabetes están directamente relacionadas al rol que juega la hiperglicemia en ésta, el cual puede ser pesquisado por las relaciones entre control glicémico y estas complicaciones. Los daños producidos por la hiperglicemia involucran complejas interacciones entre la genética del individuo, tabaquismo, índice de masa corporal, dislipidemia, alteraciones en factores de coagulación67. Los mecanismos intracelulares implicados en estas complicaciones incluyen: incremento del flujo de la vía de los polioles, activación de proteína quinasa C, incremento en la vía de las hexosaminas, y aumento de la formación de AGEs. El daño producido por estos mecanismos está relacionado al estrés oxidativo68. Muchos de los efectos de la hiperglicemia en la diabetes están mediados por los AGEs23,69-72 que llevan a la formación de intermediarios reactivos e inestables que rápidamente forman entrecruzamientos covalentes intra e intermoleculares57,73 o productos de glicoxidación74.
Estudios histopatológicos y por análisis de autofluorescencia en piel, han mostrado que, aparte de la diabetes, los AGEs se acumulan en una amplia variedad de tipos de tejidos y asociados principalmente a condiciones de inflamación crónica, incluyendo ateromas coronarios, corteza renal, membrana basal mesangial y glomerular, capa dermal, placas amiloides en enfermedad de Alzheimer, en cartílago de artritis reumatoide, músculo cardíaco, pulmón e hígado, en lupus eritematoso, osteoartritis75,76.
Por otra parte, experimentos realizados en animales demuestran que las concentraciones de AGE aumentan en aquellos que se hicieron diabéticos, y que tal incremento es sistémico en los siguientes órganos y tejidos: riñones, piel y tejido vascular77. En estudios realizados en pacientes diabéticos se ha visto que la concentración de CML aumenta en aquellos que presentan complicaciones asociadas tales como nefropatía78,79, retinopatía80 y aterosclerosis81,82. Otros estudios muestran otras alteraciones, por ejemplo, un estudio in vitro mostró que la exposición crónica de mioglobina a metil-glioxal ocasiona alteraciones en la estructura de la proteína, modificando características tales como la movilidad electroforética, contenido a-helicoidal, etc.83.
Por otra parte, la autooxidación de la glucosa va unida a la generación de especies reactivas del oxígeno tales como radicales superóxido84. Las especies reactivas del oxígeno, a su vez, aumentan la glicación68,85,86 y ambos mecanismos interfieren con una amplia variedad de procesos fisiológicos promotores de la aterogénesis87.
Aquellas proteínas de larga vida-media son más propensas a ser modificadas por la exposición a glucosa, o a derivados de ella. Entre estas proteínas, se ha estudiado que el colágeno, experimenta las siguientes modificaciones al envejecer: menor solubilidad, elasticidad y sensibilidad a digestión por proteasas, y aumento de la estabilidad térmica64. Sin embargo, todos estos cambios se ven acelerados en la diabetes88 en donde es posible identificar, por ejemplo en el colágeno, cantidades elevadas de un producto de glicación inicial (fructosil-lisina)89. Algunos ejemplos de estas proteínas son, además del colágeno, la elastina, proteínas de matriz extracelular, mielina, cartílago, cristalino, queratina y otros90,91.
Es así, como es posible aproximarnos a la edad de una proteína, mediante su grado de modificación de AGEs. La formación de AGEs puede ser una de las vías por las que nuestro cuerpo identifica aquellos blancos que están preparados para un recambio, para diferenciarlos de aquellos que han sido sintetizados más recientemente, incluso si la estructura o función son similares24.
Referencias bibliográficas
- Maillard LC. 1912. Action of amino acids on sugars. Formation of melanoidins in a methodical way. CR Acad Sci Ser 154 (2): 66-68.
- Vlassara H, Palace MR. 2003. Glycoxidation, the menace of diabetes and aging. Mt Sinai J Med 70: 232-241.
- Yamagishi S, Matsui T, Nakamura K. 2008. Possible involvement of tobacco-derived advanced glycation end products (AGEs) in an increased risk for developing cancers and cardiovascular disease in former smokers. Med Hypotheses 71 (2): 259-261.
- Bierhaus A, Hofmann MA, Ziegler R, Nawroth PP. 1998. AGE and their interaction with AGE-receptors in vascular disease and diabetes. The AGE concept. Cardiovasc Res 37: 586-600.
- Thornalley P. 2005. Dicarbonyl Intermediates in the Maillard Reaction. Ann NY Acad Sci 1043: 111-117.
- Bann H, Higgins P. 1981. Reaction of monosaccharides with proteins: possible evolutionary significance. Science 213: 222-224.
- Takagi Y, Kashiwagi A, Tanaka Y, Asahina T, Kikkawa R, Shigeta Y. 1995. Significance of fructose-induced protein oxidation and formation of advanced glycation end products. J Diab Comp 9: 87-91.
- Suárez G, Rajaram R, Oronsky A, Gawinowicz M. 1989. Nonenzymatic glycation of bovine serum albumin by fructose (fructosylation): comparision with the Maillard reaction initiated by glucose. J Biol Chem 264: 3674-3679.
- Brownlee M, Vlassara H, Cerami A. 1984. Nonenzymatic glycosylation and the pathogenesis of diabetic complications. Ann Intern Med 101: 527-537.
- Sell D, Lane M, Johnson W, Masoro E, Mock O, Reiser K, et al. 1996. Longevity and the genetic determination of collagen glycoxidation kinetics in mammalian senescence. Proc Nat Acad Sci USA 93 (1): 485-490.
- Lyons T, Silvestri G, Dunn J, Dyer D, Baynes J. 1991. Role of glycation in modification of lens crystallins in diabetic and nondiabetic senile cataracts. Diabetes 40 (8): 1010-1015.
- Tsilbary E, Charonis A, Reger L, Wohlhueter R, Furcht L. 1988. The effect of nonenzymatic glucosylation on the binding of the main noncollagenous NC1 domain to type IV collagen. J Biol Chem 263: 4302-4308.
- Charonis A, Reger L, Dege J, Kouzi-Koliakos K, Furcht L, Wohlhueter R, et al. 1990. Laminin alterations after in vitro nonenzymatic glucosylation. Diabetes 39: 807-814.
- Brownlee M, Pongor S, Cerami A. 1983. Covalent attachment of soluble proteins by nonenzymatically glycosylated collagen. J Exp Med 158: 1739-1744.
- Brownlee M, Vlassara H, Cerami A. 1985. Non-enzymatic glycosylation products on collagen covalently trap low-density lipoprotein. Diabetes 34: 938-941.
- Sensi M, Tanzi P, Bruno M, Mancuso M, Adriani D. 1986. Human glomerular basement membrane: altered binding characteristics following in vitro non-enzymatic glycosylation. Ann NY Acad Sci 488: 549-552.
- Hartoglou C, Tsilbary E, Brownlee M, Charonis A. 1992. Altered cellular interactions between endothelial cells and non-enzymatically glucosylated laminin/type IV collagen. J Biol Chem 267: 12404-12407.
- Federoff H, Lawrence D, Brownlee M. 1993. Nonenzymatic glycosylation of laminin and the laminin peptide CIKVAVS inhibits neurite outgrowth. Diabetes 42: 509-513.
- Crowley S, Brownlee M, Edelstein D, Satriano J, Mori T, Singhal P, et al. 1991. Effects of nonenzymatic glycosylation of mesangial matrix on proliferation of mesangial cells. Diabetes 40: 540-547.
- Vlassara H, Bucala R. 1996. Recent progress in advanced glycation and diabetic vascular disease: role of advanced glycation end product receptors. Diabetes 45 (S3): S65-S66.
- Singh R, Barden A, Mori T, Beilin L. 2002. Advanced glycation end-products: a review. Diabetologia 44 (2): 129-146.
- Thornalley P, Langborg A, Minhas H. 1999. Formation of glyoxal, methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose. Biochem J 344: 109-116.
- Olmos P, Araya-del-Pino A, González C, Laso P, Irribarra V, Rubio L. 2009. Fisiopatología de la retinopatía y nefropatía diabéticas. Rev Med Chile 137: 1375-1384.
- Barlovic D, Soro-Paavonen A, Jandeleit-Dahm K. 2011. RAGE biology, atherosclerosis and diabetes. Clinical Science 121: 43-55.
- Monnier VM. 1990. Nonenzymatic glycosylation, the Maillard reaction and the aing process. J Gerontol 45 (4): B105-B111.
- Uribarri J, Tuttle KR. 2006. Advanced glycation end products and nephrotoxicity of high-protein diets. Clin J Am Soc Nephrol 1: 1293-1299.
- Luevano-Contreras C, Chapman-Novakofski K. 2010. Dietary Advanced Glycation End Products and Aging. Nutrients 2: 1247-1265.
- Ahmed N. 2005. Advanced glycation end products role in pathology of diabetic complications. Diab Res Clin Pract 67: 3-21.
- Ahmed N, Thornalley PJ. 2007. Advanced glycation end products: What is their relevance to diabetic complications? Diab Obes Metab 9: 233-245.
- Sims T, Rasmussen LM, Oxlund H, Bailey AJ. 1996. The role of glycation cross-links in diabetic vascular stiffening. Diabetologia 39 (8): 946-951.
- Nowotny K, Jung T, Höhn A, Weber D, Grune T. 2015. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 5 (1): 194-222.
- Monnier VM, Sell D, Genuth S. 2005. Glycation products as markers and predictors of the progression of diabetic complications. Ann NY Acad Sci 1043: 567-581.
- Goldin A, Beckman JA, Schmidt AM. 2006. Advanced glycation end products: sparking the development of diabetic vascular injury. Circulation 114 (6): 597-605.
- Genuth S, Sun W, Cleary P, Sell D, Dahms W, Malone J, et al. 2005. Glycation and carboxymethyllysine levels in skin collagen predict the risk of future 10-year progression of diabetic retinopathy and nephropathy in the diabetes control and complications trial and epidemiology of diabetes interventions and complications participants with type 1 diabetes. Diabetes 54 (11): 3103-3111.
- American Diabetes Association. 2013. Standards of medical care in diabetes. Diabetes Care 36 (S1): S4-S10.
- Ferrer S. Diabetes y neurología, capítulo Bioquímica de la diabetes mellitus (páginas 13-37), Pablo Olmos. Editorial Iku. Chile, 2008.
- Schmidt AM, Vianna M, Gerlach M, Brett J, Ryan J, Kao J, et al. 1992. Isolation and Characterization of Two Binding Proteins for Advanced Glycosylation End Products from Bovine Lung Which Are Present on the Endothelial Cell Surface. J Biol Chem 267: 14987-14997.
- Neeper M, Schmidt AM, Brett J, Yan SD, Wang F, Pan YC, et al. 1992. Cloning and expression of a cell surface receptor for advanced glycosylation end products of proteins. J Biol Chem 267: 14998-5004.
- Sakurai S, Yonekura H, Yamamoto Y, Watanabe T, Tanaka N, Li H, et al. 2003 The AGE-RAGE system and diabetic nephropathy. J Am Soc Nephrol 14 (S3): S259-S263.
- Yonekura H, Yamamoto Y, Sakurai S, Petrova RG, Abedin MJ, Li H, et al. 2003. Novel splice variants of the receptor for advanced glycation end-products expressed in human vascular endothelial cells and pericytes, and their putative roles in diabetes-induced vascular injury. Biochem J 370: 1097-1109.
- Schmidt A, Stern D. 2001. Receptor for age (RAGE) is a gene within the major histocompatibility class III region: implications for host response mechanisms in homeostasis and chronic disease. Front Biosci 6: D1151-D1160.
- Bierhaus A, Nawroth PP. 2009. Multiple levels of regulation determine the role of the receptor for AGE (RAGE) as common soil in inflammation, immune responses and diabetes mellitus and its complications. Diabetologia 52 (11): 2251-2263.
- Bierhaus A, Humpert P, Morcos M, Wendt T, Chavakis T, Arnold B, et al. 2005. Understanding RAGE, the receptor for advanced glycation end products. J Mol Med 83: 876-886.
- Yan SD, Schmidt AM, Anderson GM, Zhang J, Brett J, Zou YS, et al. 1994. Enhanced cellular oxidant stress by the interaction of advanced glycation end products with their receptors/binding proteins. J Biol Chem 269 (13): 9889-9897.
- Huttunen HJ, Kuja-Panula J, Rauvala H. 2002. Receptor for advanced glycation end products (RAGE) signaling induces CREB-dependent chromogranin expresión during neural differentiation. J Biol Chem 277 (4): 38635-38646.
- Chang JS, Wendt T, Qu W, Kong L, Zou YS, Schmidt AM, et al. 2008. Oxygen deprivation triggers upregulation of early growth response-1 by the receptor for advanced glycation end products. Xirc Res 102 (8): 905-913.
- Zeng S, Feirt N, Goldstein M, Guarrera J, Ippagunta N, Ekong U, et al. 2004. Blockade of receptor for advanced glycation endproducts (RAGE) attenuates ischemia and reperfusion injury in the liver in mice. Hepatology 39 (2): 422-432.
- Taguchi A, Blood DC, del Toro G, Canet A, Lee DC, Qu W, et al. 2000. Blockade of RAGE-amphoterin signalling suppresses tumour growth and metastases. Nature 405 (6784): 354-360.
- Lander HM, Tauras JM, Ogiste JS, Hori O, Moss RA, Schmidt AM. 1997. Activation of the receptor for advanced glycation endproducts triggers a p21(ras)-dependent mitogen-activated protein kinase pathway regulated by oxidant stress. J Biol Chem 272 (28): 17810-17814.
- Marsche G, Semlitsch M, Hammer A, Frank S, Weigle B, Demling N, et al. 2007. Hypochlorite-modified albumin colocalizes with RAGE in the artery wall and promotes MCP-1 expression via the RAGE-ERK 1/2 MAP-kinase pathway. FASEB J 21 (4): 1145-1152.
- Reddy MA, Lis SL, Sahar S, Kim YS, Xu ZG, Lanting L, et al. 2006. Key role of Src kinase in S100b-induced activation of the receptor for advanced glycation endproducts in vascular smooth muscle cells. J Biol Chem 281 (19): 13685-13693.
- Sakaguchi T, Yan SF, Yan SD, Belov D, Rong LL, Sousa M, et al. 2003. Central role of RAGE-dependent neointimal expansion in arterial restenosis. J Clin Invest 111 (7): 959-972.
- Li YM, Mitsuhashi T, Wojciechowicz D, Shimizu N, Li J, Stitt A, et al. 1996. Molecular identity and cellular distribution of advanced glycation endproduct receptors: relationship of p60 to OST-48 and p90-80K-H membrane proteins. Proc Natl Acad Sci USA 93: 11047-11052.
- Vlassara H, Li YM, Imani F, Wojciechowicz D, Yang Z, Liu F, et al. 1995. Identi fication of galectin-3 as a high-af finity binding protein for advanced glycation end products (AGE): a new member of the AGE-receptor complex. Mol Med 1: 634-646.
- Stitt AW, He C, Vlassara H. 1999. Characterization of the advanced glycation end-product receptor complex in human vascular endothelial cells. Biochem Biophy Res Commun 256: 549-556.
- Stitt AW, Li YM, Gardiner TA, Bucala R, Archer DB, Vlassara H. 1997. Advanced glycation end products (AGEs) co-localize with AGE receptors in the retinal vasculature of diabetic and of AGE-infused rats. Am J Pathol 150: 523-531.
- Koschinsky T, He CJ, Mitsuhashi T, Bucala R, Liu C, Buenting C, et al. 1997. Orally absorbed reactive glycation products (glycotoxins): An environmental risk factor in diabetic nephropathy. Proc Natl Acad Sci USA 94: 6474-6479.
- Uribarri J, Woodruff S, Goodman S, Cai W, Chen X, Pyzik R, et al. 2010. Advanced glycation end products in foods and a practical guide to their reduction in the diet. J Am Diet Assoc 110 (6): 911-16.e12.
- Makita Z, Radoff S, Rayfield EJ. 1991. Advanced glycosilation end products in diabetic nephropathy. New Eng J Med 325: 836-842.
- Makita Z, Bucala R, Rayfield EJ, Friedman EA, Kaufman AM, Korbet SM, et al. 1994. Reactive glycosylation end products in diabetic uraemia and treatment of renal failure. Lancet 343 (8912): 1519-1522.
- Bucala R, Makita Z, Vega G, Grundy S, Koschinsky T, Cerami A, et al. 1994. Modification of low density lipoprotein by advanced glycation end products contributes to the dyslipidemia of diabetes and renal insufficiency. Proc Natl Acad Sci USA 91 (20): 9441-9445.
- O’Brien J, Morrissey PA. 1989. Nutritional and toxicological aspects of the Maillard browning reaction in foods. Crit Rev Food Sci Nutri 28 (3): 211-248.
- Finot PA, Magnenat E. 1981. Metabolic transit of early and advanced Maillard products. Prog Food Nutr Sci 5 (1-6): 193-207.
- McCance D, Dyer D, Dunn J, Bailie K, Thorpe S, Baynes J, et al. 1993. Maillard reaction products and their relation to complications in insulin-dependant diabetes mellitus. J Clin Invest 91 (6): 2470-2478.
- Vlassara H, Fuh H, Makita Z, Krungkrai S, Cerami A, Bucala R. 1992. Exogenous advanced glycosylation end products induce complex vascular dysfunction in normal animals: a model for diabetic and aging complications. Proc Nat Acad Sci USA 89 (24): 12043-12047.
- Vitek M, Bhattacharya K, Glendening JM, Stopa E, Vlassara H, Bucala R, et al. 1994. Advanced glycation end products contribuye to amyloidosis in Alzheimer disease. Proc Natl Acad Sci USA 91 (11): 4766-4770.
- Farouque HM, O’Brien RC, Meredith IT. 2000. Diabetes and coronary heart disease-from prevention to intervention: Part 1. Aust N Z J Med 30: 351-359.
- Brownlee M. 2001. Biochemistry and molecular cell biology of diabetic complications. Nature 414: 813-820.
- Vlassara H, Bucala R, Striker L. 1994. Pathogenic effects of advanced glycosylation: biochemical, biologic, and clinical implications for diabetes and aging. Lab Invest 70 (2): 138-151.
- Vlassara H, Palace MR. 2002. Diabetes and advanced glycation end products. J Intern Med 251: 87-101.
- Brownlee M, Cerami A, Vlassara H. 1988. Advanced glycosylation end products in tissue and the biochemical basis of diabetic complications. N Engl J Med 318 (20): 1315-1321.
- Olmos P, Niklitschek S, Olmos R, Faúndez J, Quezada T, Bozinovic M, et al. 2012. Bases fisiopatológicas para una clasificación de la neuropatía diabética. Rev Med Chile 140: 1593-1605.
- Grandhee SK, Monnier VM. 1991. Mechanism of formation of the Maillard protein cross-link pentosidine. Glucose, fructose, and ascorbate as pentosidine precursors. J Biol Chem 266 (18): 11649-11653.
- Wells-Knecht KJ, Zyzak DV, Litchfield JE, Thorpe SR, Baynes JW. 1995. Mechanism of autoxidatuve glycosylation: identification of glyoxal and arabinose as intermediates in the autoxidative modification of protein by glucose. Biochemistry 34 (11): 3702-3709.
- Matsumoto T, Tsurumoto T, Baba H, Osaki M, Enomoto H, Yonekura A, et al. 2007. Measurement Measurement of advanced glycation endproducts in skin of patients with rheumatoid arthritis, osteoarthritis, and dialysis-related spondyloarthropathy using non-invasive methods. Rheumatol Int 28 (2): 157-160.
- De Leeuw K, Graaff R, De Vries R, Dullaart RP, Smit AJ, Kallenberg CG, et al. 2007. Accumulation of advanced glycation end products in patients with systemic lupus erythematous. Rheumatology 46 (10): 1551-1556.
- Baynes JW, Thorpe SR. 1999. Role of oxidative stress in diabetic complications a new perspective on an old paradigm. Diabetes 48: 1-9.
- Makino H, Shikata K, Hironaka K, Kushiro M, Yamasaki Y, Sugimoto H, et al. 1995. Ultrastructure of nonenzymatically glycated mesangial matrix in diabetic nephropathy. Kidney Int 48: 517-526.
- Suzuki D, Yagame M, Jinde K, Naka R, Yano N, Endoh M, et al. 1996. Immunofluorescence staining of renal biopsy samples in patients with diabetic nephropathy in non-insulin-dependent diabetes mellitus using monoclonal antibody to reduced glycated lysine. J Diabetes Complications 10: 314-319.
- Murata T, Nagai R, Ishibashi T, Inomuta H, Ikeda K, Horiuchi S. 1997. The relationship between accumulation of advanced glycation end products and expression of vascular endothelial growth factor in human diabetic retinas. Diabetologia 40: 764-769.
- Kume S, Takeya M, Mori T, Araki N, Suzuki H, Horiuchi S, et al. 1995. Immunohistochemical and ultrastructural detection of advanced glycation end products in atherosclerotic lesions of human aorta with a novel specific monoclonal antibody. Am J Pathol 147: 654-667.
- Sakata N, Imanaga Y, Meng J, Tachikawa Y, Takebayashi S, Nagai R, et al. 1999. Increased advanced glycation end products in atherosclerotic lesions of patients with end-stage renal disease. Atherosclerosis 142: 67-77.
- Banerjee S, Chakraborti AS. 2013. In viro study on structural alteration of myoglobin by methylglyoxal. Protein J 32 (3): 216-222.
- Wolff SP, Dean RT. 1987. Glucose autoxidation and protein modification. The potential role of “autoxidative glycosylation” in diabetes. Biochem J 245 (1): 243-250.
- Nishikawa T, Edelstein D, Du XL, Yamagishi S, Matsumura T, Kaneda Y, et al. 2000. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature 404 (6779): 787-790.
- Vlassara H, Uribarri J. 2004. Glycoxidation and diabetic complications: modern lessons and a warning? Rev End & Metab Dis 5: 181-188.
- Bucala R, Mitchell R, Arnold K, Innerarity T, Vlassara H, Cerami A. 1995. Identification of the major site of apolipoprotein B modification by advanced glycosylation end products blocking uptake by the low density lipoprotein receptor. J Biol Chem 270 (18): 10828-10832.
- Hamlin CR, Kohn RR, Luschin JH. 1975. Apparent accelerated aging of human collagen in diabetes mellitus. Diabetes 24: 902-904.
- Dyer DG, Dunn JA, Thorpe SR, Lyons TJ, McCance DR, Baynes JW. 1993. Accumulation of Maillard reaction products in skin collagen in diabetes and aging. J Clin Invest 91: 2463-2469.
- Fu M, Wells-Knecht K, Blackledge J, Lions T, Thorpe S, Baynes J. 1994. Glycation glycoxidation and crosslinking of collagen by glucose: kinetics, mechanisms and inhibition of late stages of the Maillard reaction. Diabetes 43: 676-683.
- Garlick R, Mazer J, Higgins P, Bun H. 1983. Characterization of glycosylated hemoglobins. Relevance to monitoring of diabetic control and analysis of other proteins. J Clin Invest 71 (5): 1062-1072.